The Makino Lab
Research
The rate of cGMP synthesis modulates visual transduction
In retinal rods, cGMP is the second messenger that links photon capture by rhodopsin on internal disk membranes to ion channel activity on the plasma membrane. Rhodopsin photoexcitation leads to the hydrolysis of cGMP and subsequent closure of cGMP-gated channels, curtailing the entry of Na+ and Ca2+. During response recovery, retina-specific guanylyl cyclases (retGCs) replenish cGMP, reopen the channels, and restore the influx of cations. To facilitate the recovery, guanylyl cyclase activating proteins (GCAPs) sense the decrease in Ca2+ caused by illumination and greatly stimulate the rate of cGMP synthesis.
All vertebrate rods use at least two GCAPs: GCAP1 and GCAP2. Why are 2 types of GCAPS necessary? Working closely with the Dizhoor laboratory at Salus University, we explored the basis for the dual system by using electrophysiological methods to study rods of mutant mice that lack one or both GCAPs. Deletion of GCAP2 did not change the amplitude of the single photon response, but slowed its recovery (Fig. 1). Elimination of GCAP1 caused the photon response to rise for twice as long to an amplitude that was twice as large. Although knockout of GCAP2 did not affect GCAP1 expression, knockout of GCAP1 did cause an up-regulation of GCAP2 as detected by immunofluorescence and Western blot. The overexpression of GCAP2 resulted in acceleration of the response recovery rather than the slowdown that was expected from the loss of GCAP1 alone.
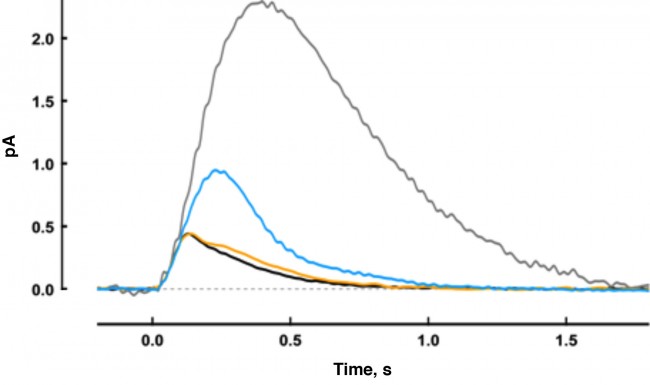
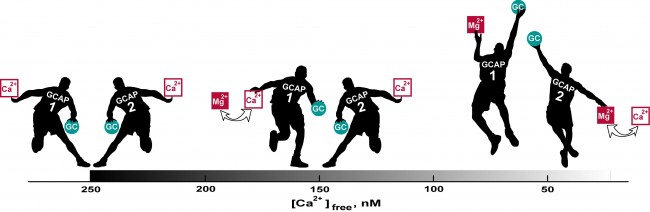
Thus the two GCAPs regulate retGCs sequentially (Fig. 2). Because of its lower Ca2+ affinity, GCAP1 is the first responder and acts to limit response amplitude. GCAP2, with a higher Ca2+ affinity, does not assist retGCs until Ca2+ concentration declines even further, which happens somewhat during a single photon response, but to a greater extent with bright light. GCAP2 stimulation of retGCs provides for a faster response recovery. Together, the two GCAPs tune the operating range and temporal resolution of rod photoreceptors.
The Sharma and Duda laboratories at Salus University discovered that bicarbonate ions stimulate cGMP synthesis by retGC, independent of GCAPs. In single cell recordings, we discovered that bicarbonate operated synergistically with GCAPs and low Ca2+ in rods and in cones. As a result, bicarbonate boosted the size of the maximal flash response, reduced relative sensitivity to flashes and altered response kinetics (Fig. 3).
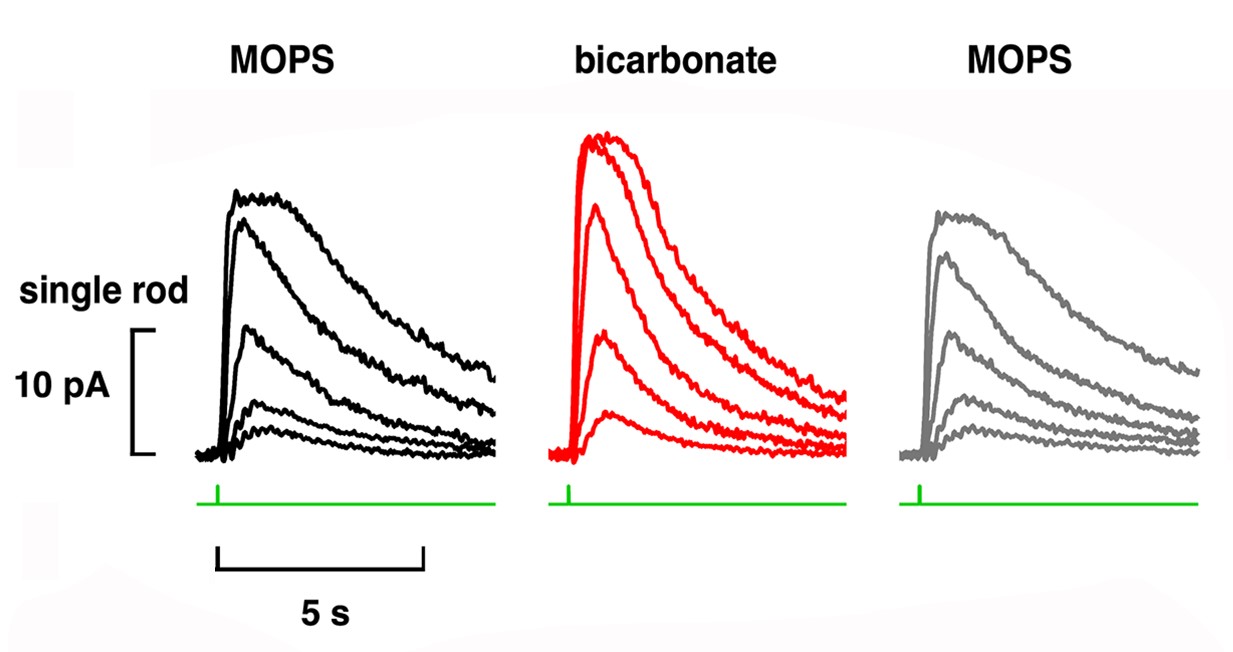
Rods do not express carbonic anhydrase, suggesting that modulation of visual transduction involves bicarbonate produced exogenously. We found that only rods retaining an intact spherule were affected by bicarbonate, indicating that uptake occurred in the inner segment, near the synaptic ending. In contrast, cones took up bicarbonate at their outer segments as well as at their inner segments. Moreover, cones did express carbonic anhydrase, which meant that they were affected by endogenous as well as exogenous bicarbonate.
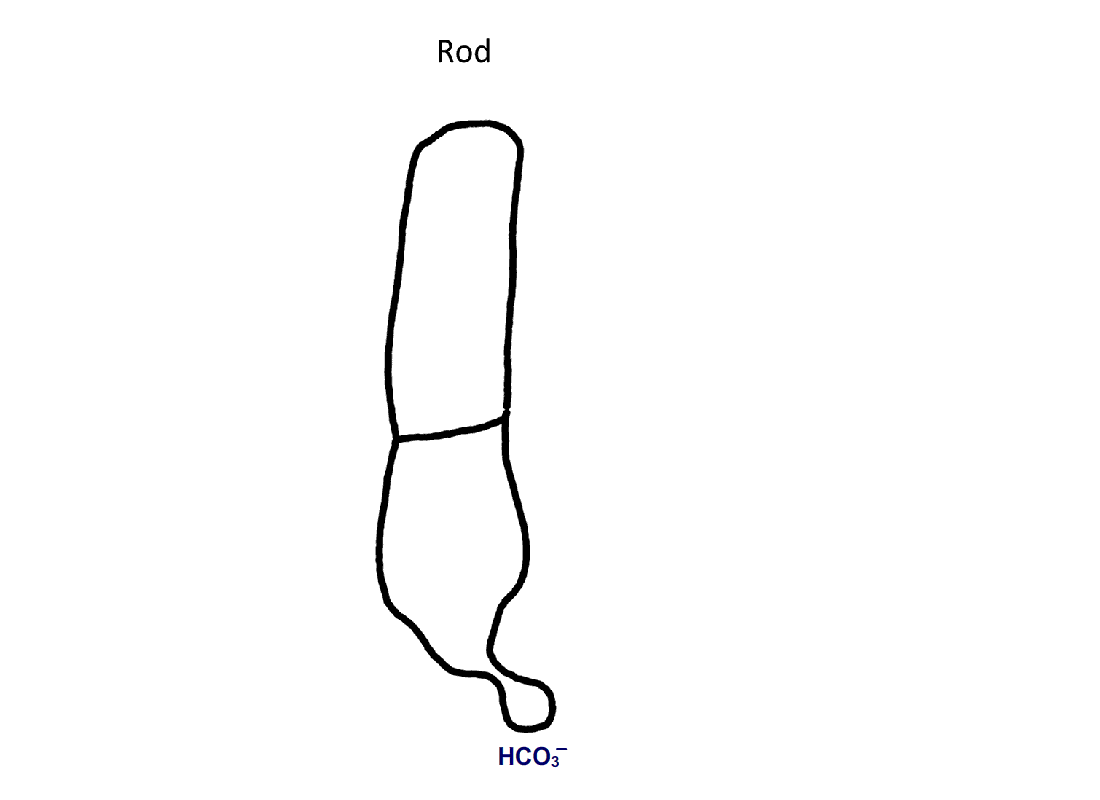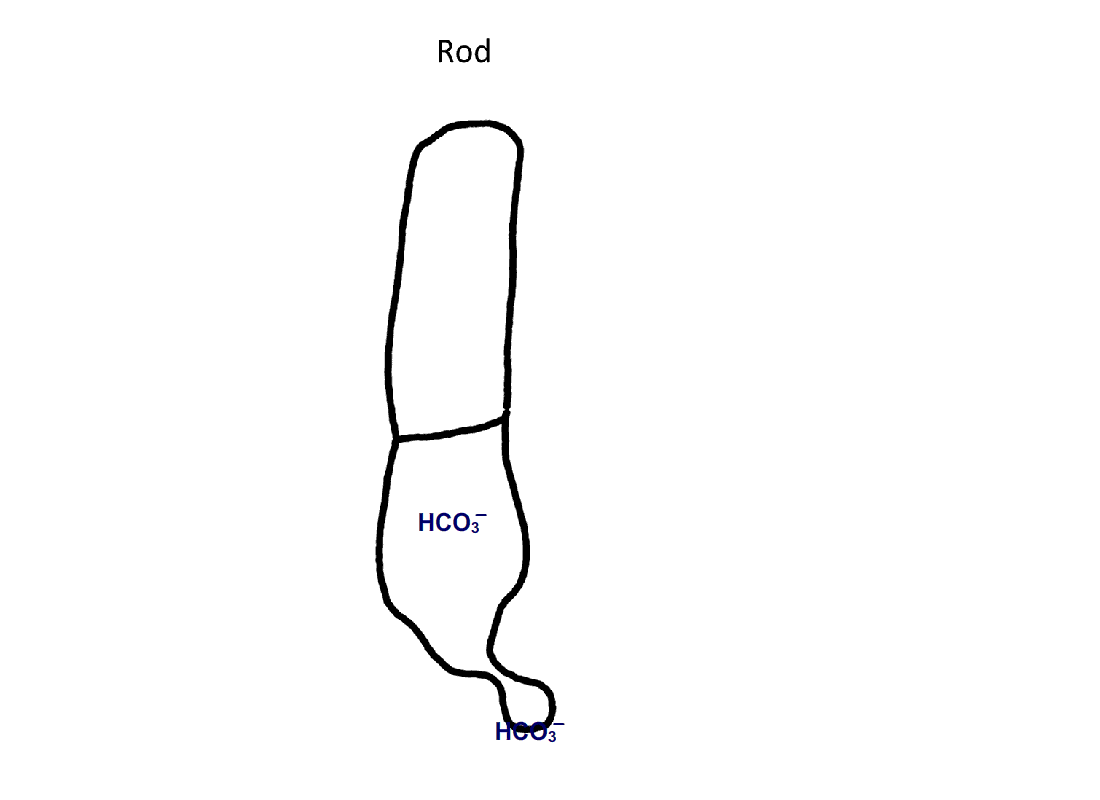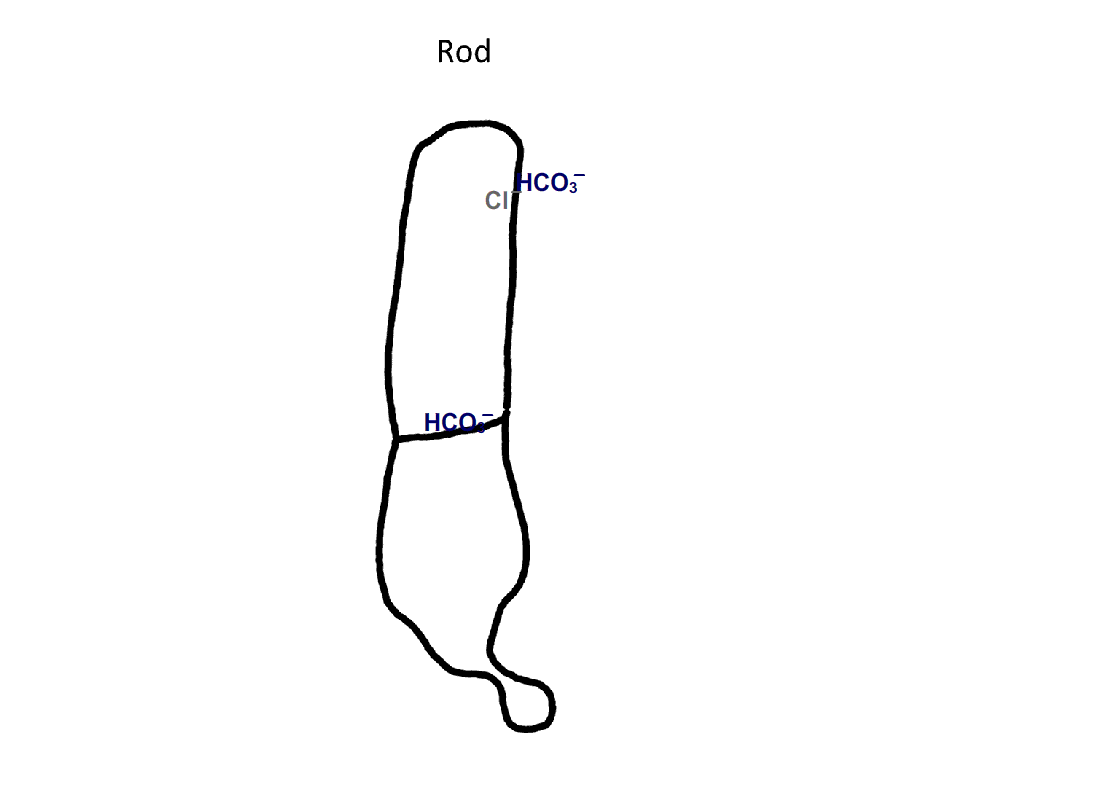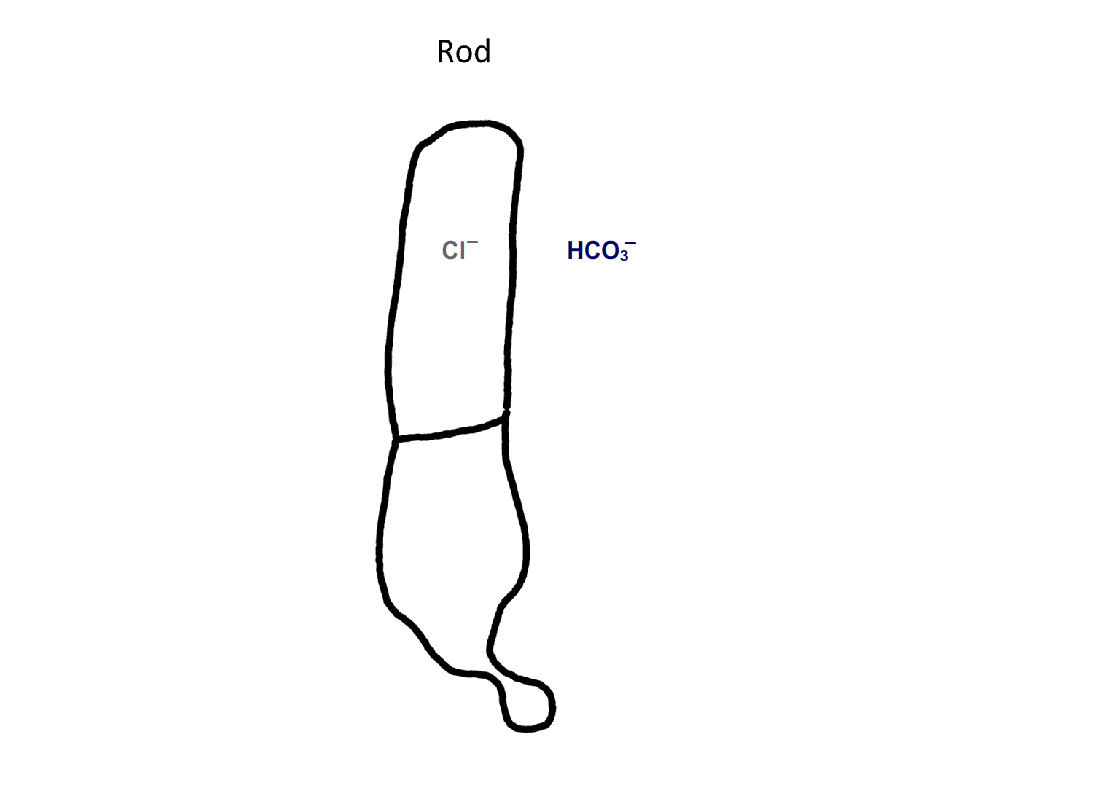
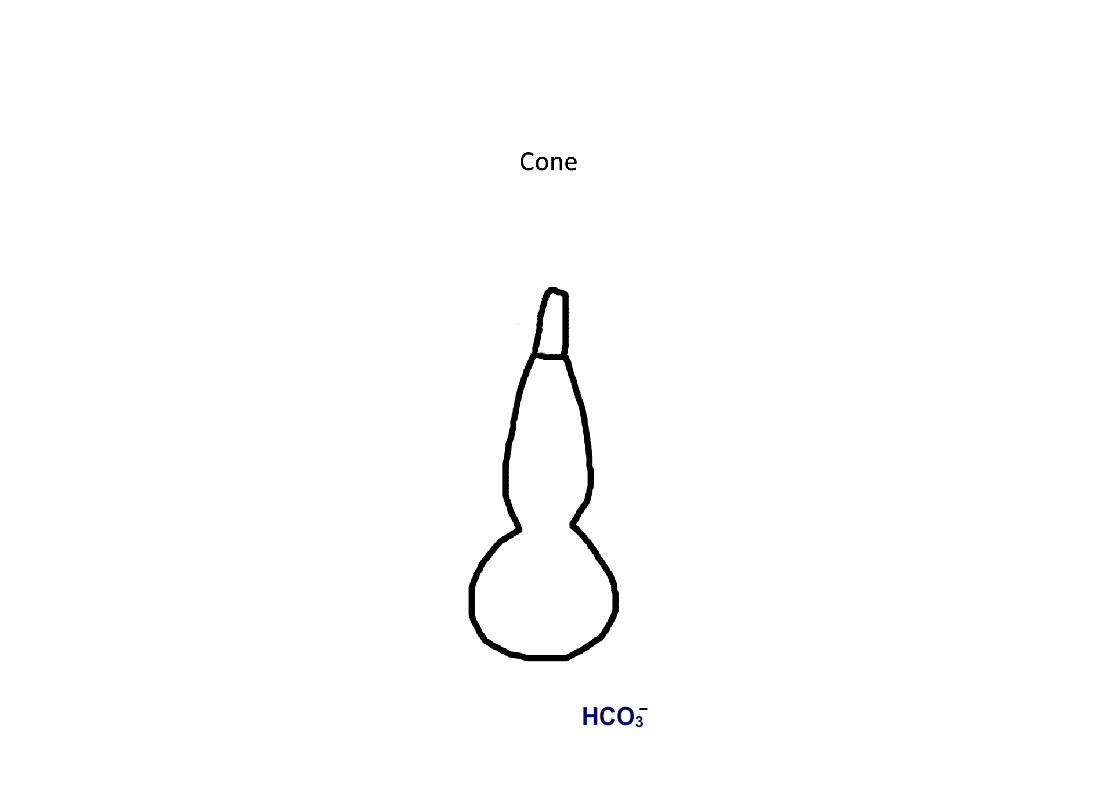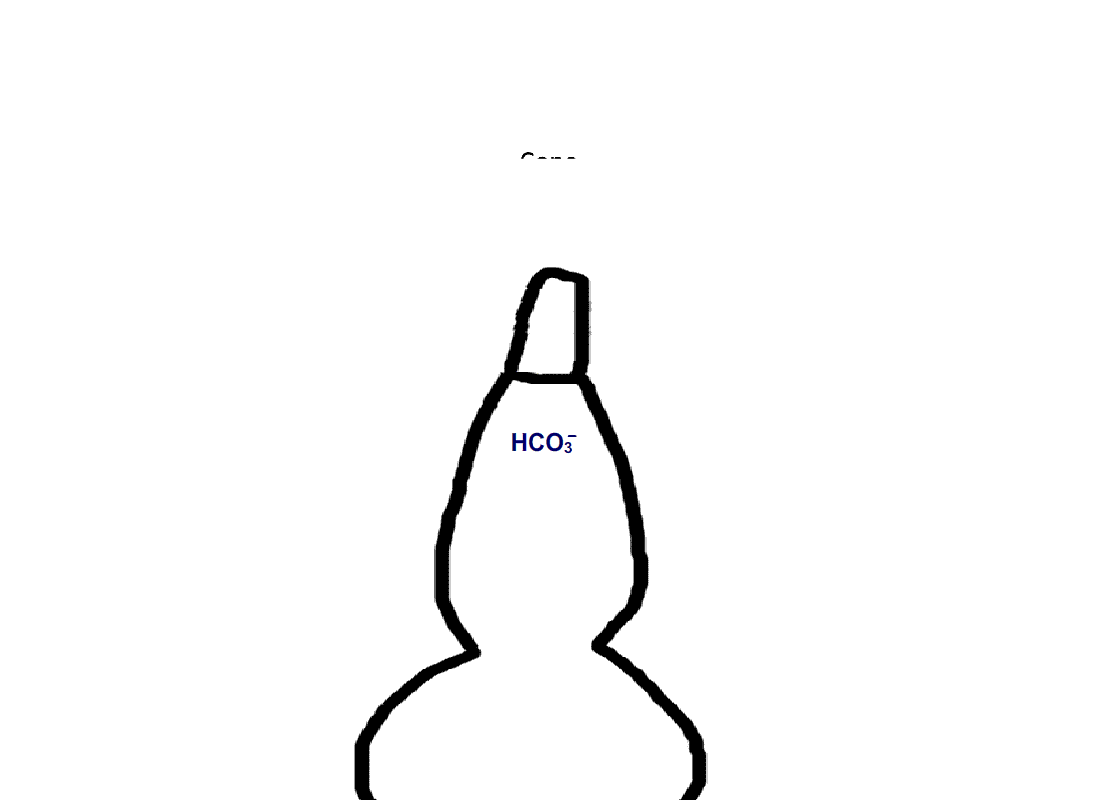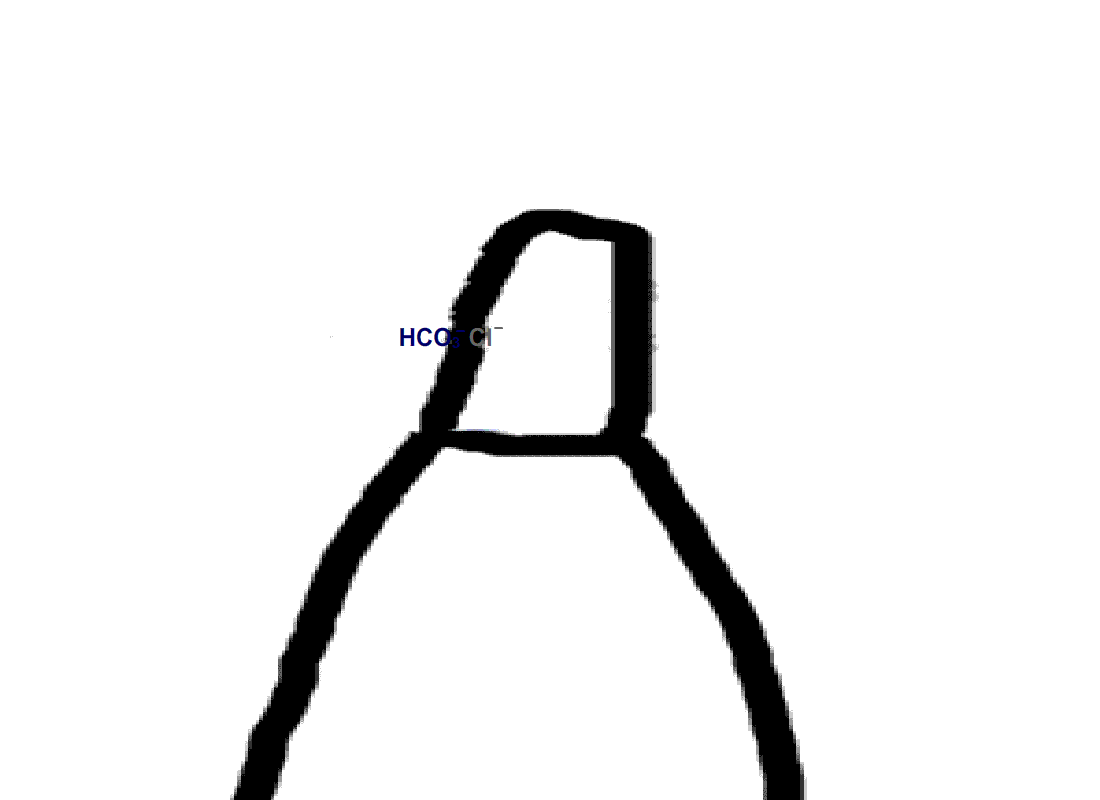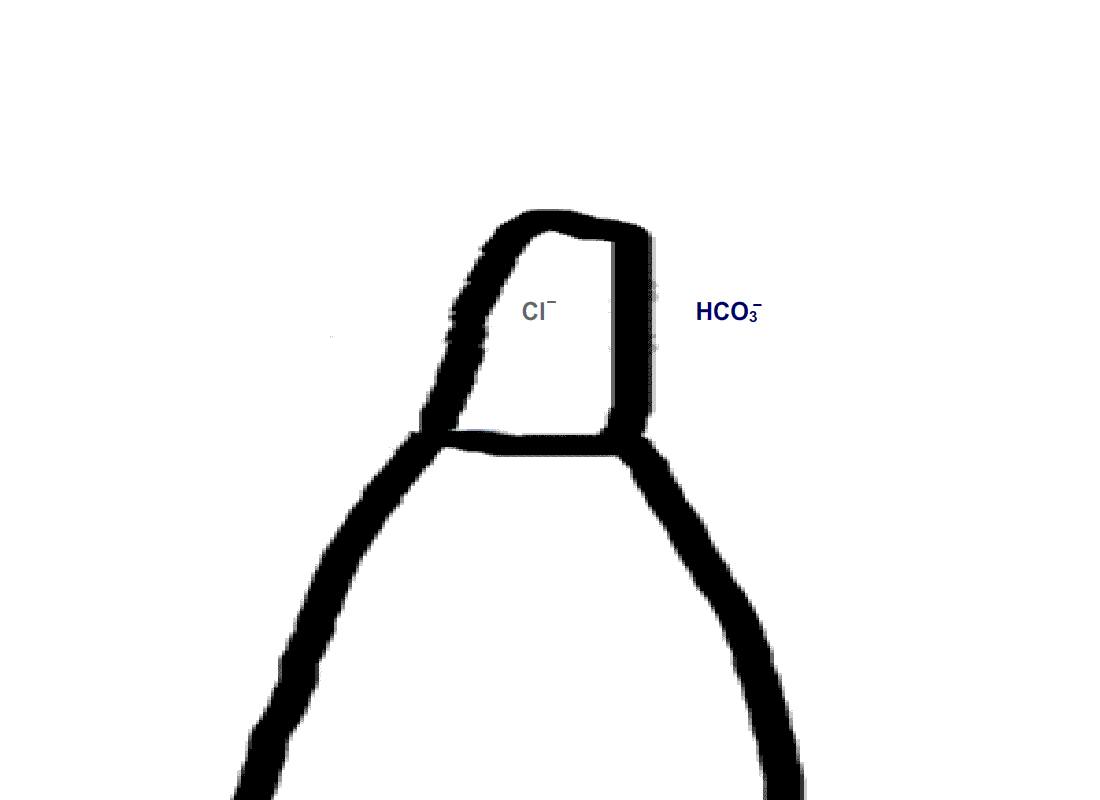
Figure 4. Pathway for bicarbonate through rods and cones. Bicarbonate enters rods and cones at their synapses (1). Cones are also able to take up bicarbonate at their outer segments, perhaps by reversed action of a bicarbonate/Cl- exchanger (2). Carbon dioxide is able to pass through the plasma membrane and in cones, but not in rods, gets converted to bicarbonate by carbonic anhydrase. Bicarbonate is removed from rods and cones by a bicarbonate/Cl- exchanger located in their outer segments.
Recent Publications
Full listing of citations on PubMed
PAPERS
Makino, C.L., Talukder, Md. S.U., Kim, B., Adhikari, R.D., Hamm, H.E., Gurevich, V.V., Klaus, C. and Caruso, G. (2025) The effects of bicarbonate on the aberrant photon response in murine rod photoreceptors. Biophys J, in press. doi: 10.1016/j.bpj.2025.06.035.
Adhikari, R.D., Kossoff, A.M., Cornwall, M.C. and Makino, C.L. (2023) Bicarbonate boosts flash response amplitude to augment absolute sensitivity and extend dynamic range in murine retinal rods. Front Mol Neurosci, 16: 1125006. PMCID: PMC10140344.
Caruso, G., Klaus, C., Hamm, H.E., Gurevich, V.V., Bisegna, P., Andreucci, D., DiBenedetto, E. and Makino, C.L. (2023) Pepperberg plot: Modeling flash response saturation in retinal rods of mouse. Front Mol Neurosci, 15: 1054449. PMCID: PMC9880052.
Geva, P., Caruso, G., Klaus, C.J., Hamm, H.E., Gurevich, V.V., DiBenedetto, E. and Makino, C.L. (2022) Effects of cell size and bicarbonate on single photon response variability in retinal rods. Front Mol Neurosci, 15: 1050545. PMCID: PMC9796569.
Klaus, C., Caruso, G., Gurevich, V.V., Hamm, H.E., Makino, C.L. and DiBenedetto, E. (2021) Phototransduction in retinal cones: Analysis of parameter importance. PLoS One, 16: e0258721. PMCID: PMC8553137.
Caruso, G., Klaus, C., Hamm, H.E., Gurevich, V.V., Makino, C.L. and DiBenedetto, E. (2020) Position of rhodopsin photoisomerization on the disk surface confers variability to the rising phase of the single photon response in vertebrate rod photoreceptors. PLoS One, 15: e0240527. PMCID: PMC7556485.
BOOK CHAPTERS
MacLeish, P.R. and Makino, C.L. (2024) Photoresponses of rods and cones, in Adler’s Physiology of the Eye 12th edition, eds. L.A. Levin, P.L. Kaufman and M.E. Hartnett. Elsevier, pp. 432-450.
TUTORIAL
Horrigan, D and Makino, C.L. (2022) How do we see? An introduction to the biophysics of visual transduction. Biophysical Society Website:
https://www.biophysics.org/Portals/0/BPSAssets/Articles/An_introduction_to_the_biophysics_of_visual_transductionv5_1.pdf
Lab Members

Andrew Best

Aanvi Trivedi
Past Lab Members:
Md S Uddin Talukder, Pd.D.
Brianna Kim
Rajan Adhikari, Ph.D.
Anaid Perez
Polina Geva, Ph.D.
Amanda Kossoff
Paris Chakravarty
Links:
Contact Us
Department of Pharmacology, Physiology & Biophysics
Chobanian & Avedisian School of Medicine
700 Albany Street W402B
Boston, MA 02118-2526
Phone: (617) 358-8470
e-mail: cmakino@bu.edu